

Cuore
La muscolatura cardiaca si contrae e si rilascia alternativamente nel corso di un ciclo cardiaco.
Cellula miocardica funzionale: possiede un citoplasma differenziato in tubuli e vescicole, delimitati da membrane che costituiscono il sistema sarcotubulare, e in strutture fibrillari contrattili.
Si distinguono quindi 3 parti:- un ambiente esterno alla cellula, in cui è presente il LEC;
- un ambiente interno alla cellula, contenente il LIC, con ioni e altre sostanze;
- sistema miofibrillare con mitocondri, che insieme ad una continua spesa energetica di ATP, mantengono le differenze di concentrazione ionica tra interno ed esterno della cellula. Ciò sta quindi alla base della differenza di potenziale elettrico della cellula miocardica, che a riposo è circa -80mV (interno negativo rispetto all’esterno).
Per potersi contrarre, la cellula miocardica deve sviluppare un p.a., quindi deve andare incontro ad una serie rapida di eventi che portino il potenziale di membrana a valori sempre più bassi (depolarizzazione rapida) e lo mantengano per qualche tempo (plateau), finchè non sarà ripristinato il valore antecedente al p.a. (ripolarizzazione). Nelle cellule nervose, questi eventi dipendono dalle variazioni delle conduttanze di membrana per gli ioni Na e K, mentre nelle cellule cardiache si aggiunge anche lo ione Ca, responsabile della fase di plateau.
- Le cellule miocardiche che avviano la contrazione sono raggruppate nel nodo del seno (SA) situato nell’atrio dx.
- Da qui, l’impulso viene trasmesso, attraverso tre fasci, ad una struttura di raccolta situata nel setto interatriale, detta nodo atrioventricolare (AV).
- Da qui poi lo stimolo è trasmesso alle restanti parti del miocardio tramite un sistema di conduzione costituito dal fascio di His, che si suddivide in due branche, le quali terminano nelle fibre di Purkinje, che a loro volta contattano le cellule della muscolatura ventricolare.
È importante anche la successione cronologica di questi eventi: il p.a. infatti, viene trasmesso dalle diverse vie di conduzione con dei ritardi. L’impulso giunge poi all’interno delle singole miocellule tramite il sistema di tubuli T, e si propaga da una cellula all’altra grazie ai dischi intercalari, che consentono sia la coesione meccanica, che la connessione elettrica, agendo quindi come sinapsi elettriche (differenza con il muscolo scheletrico, in cui le unità motorie sono attivate attraverso impulsi nervosi trasmessi a livello della placca motrice, che è invece una sinapsi chimica).
ruota la testa consentendo lo scorrimento dei filamenti e quindi la contrazione del miocardio.
Dopo la contrazione, si ripristina lo stato iniziale attraverso la ripolarizzazione, e il Ca viene riportato nel reticolo sarcoplasmatico.
- la fase di depolarizzazione rapida dura circa 2ms, determinata da un forte incremento della permeabilità al Na;- la fase di plateau dura circa 200ms, ed è dovuta all’incremento della permeabilità al Ca, mentre allo stesso tempo si ha una diminuzione della permeabilità al K;
- i due periodi precedenti costituiscono il periodo refrattario assoluto della cellula, che è poi seguito da un periodo refrattario relativo di circa 50ms.
La risposta meccanica inizia con gradualità subito dopo la depolarizz. rapida, raggiunge il suo max verso la fine del plateau e si riduce fino ad azzerarsi circa 50ms dopo la fine del p.a.
La lunga durata del p.a. e la lunga durata del periodo refrattario assoluto fanno sì che non si verifichino condizioni di tetano muscolare (a differenza del muscolo scheletrico).
Cellule del nodo SA: sono dette pacemaker, in quanto avviano l’attività cardiaca.
Queste cellule non hanno un potenziale di riposo stabile, ma tendono spontaneamente alla depolarizzazione fino al raggiungimento del valore di soglia per la generazione del p.a., il quale è diverso da quello delle cellule funzionali del miocardio, in quanto non presenta la rapida depolarizzazione e la fase di plateau. Il p.a. delle cellule pacemaker quindi ha una pendenza positiva, che porterebbe il nodo SA ad una frequenza di scarica intrinseca pari a 100/110 battiti al min: in realtà però, dato che la frequenza a riposo va da un min di 70 ad un max variabile con l’età, esiste un meccanismo di regolazione che consente alla frequenza a riposo di stabilizzarsi su un valore inferiore a quello intrinseco, e di elevarsi, quando necessario, anche più di tre volte sopra al valore di riposo. Questo meccanismo è regolato dal SNA, che innerva il cuore con la branca simpatica e parasimpatica, con un’azione cronotropa (regolatrice della frequenza), e un’azione inotropa (regolatrice della forza di contrazione):
- il sistema simpatico innerva atri e ventricoli e garantisce un’azione cronotropa positiva e inotropa positiva;
- il sistema parasimpatico innerva il nodo SA e il nodo AV e garantisce un’azione cronotropa negativa.
Le terminazioni dei due sistemi si suddividono poi tra le varie fibrocellule muscolari, le quali possiedono sia recettori adrenergici per la noradrenalina del simpatico, sia recettori muscarinici per l’Ach del parasimpatico.
Ciclo cardiaco: la serie di eventi elettrici e meccanici che consentono al cuore di svolgere la sua attività di pompa sono descritti secondo la curva pressione-volume dei ventricoli e il diagramma di Wiggers.
Curva pressione-volume del ventricolo sinistro:
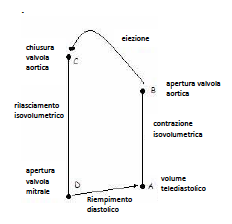
• A: fine del riempimento del ventricolo, con volume telediastolico , (circa 130ml di sangue). In seguito inizia la sistole, la valvola mitrale si chiude per impedire rigurgiti di sangue e anche la valvola aortica è chiusa;
• A-B: si ha un continuo aumento della pressione ventricolare mantenendo sempre lo stesso volume, quindi si ha una contrazione isovolumetrica;
• B: a questo punto, la pressione ventricolare supera quella in aorta (80mmHg) e ciò fa aprire la valvola aortica in cui viene spinto il sangue, con la fase di eiezione (B-C). In questa fase, il ventricolo raggiunge la max pressione, di circa 120mmHg, ed espelle un volume di sangue di circa 70-90ml (65% del volume telediastolico);
• C: finisce l’eiezione ventricolare, la valvola aortica si chiude, la pressione cala e inizia la diastole, con il rilasciamento isovolumetrico;
• D: a questo punto il volume ventricolare è minimo, con rimanenti circa 50ml di sangue, ed è detto telesistolico. Da questo ricomincia poi un nuovo ciclo, con la riapertura della valvola mitrale, permettendo al sangue di passare dall’atrio al ventricolo.
• A-B: si ha un continuo aumento della pressione ventricolare mantenendo sempre lo stesso volume, quindi si ha una contrazione isovolumetrica;
• B: a questo punto, la pressione ventricolare supera quella in aorta (80mmHg) e ciò fa aprire la valvola aortica in cui viene spinto il sangue, con la fase di eiezione (B-C). In questa fase, il ventricolo raggiunge la max pressione, di circa 120mmHg, ed espelle un volume di sangue di circa 70-90ml (65% del volume telediastolico);
• C: finisce l’eiezione ventricolare, la valvola aortica si chiude, la pressione cala e inizia la diastole, con il rilasciamento isovolumetrico;
• D: a questo punto il volume ventricolare è minimo, con rimanenti circa 50ml di sangue, ed è detto telesistolico. Da questo ricomincia poi un nuovo ciclo, con la riapertura della valvola mitrale, permettendo al sangue di passare dall’atrio al ventricolo.
Il cuore deve lavorare contro due carichi:
- precarico, in quanto man mano che il cuore si riempie, le fibre si stirano e la massa di sangue rappresenta il carico applicato al muscolo prima che inizi la contrazione. Esiste un valore ottimale di precarico da cui il muscolo cardiaco è in grado di sviluppare la maggior forza contrattile;
- postcarico, rappresentato dalla pressione diastolica dell’aorta e delle arterie polmonari che il cuore deve vincere per potervi spingere il sangue.
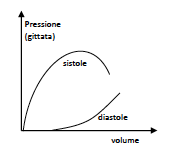
Il cuore ha la capacità intrinseca di adattarsi a variazioni del volume di sangue venoso in ingresso: infatti, più aumenta il volume telediastolico, più le fibre si stirano e più aumentano la forza di contrazione e la gittata. Oltre un certo volume telediastolico però, la gittata diminuisce, indicando
quindi che il ventricolo è troppo dilatato ed è diminuita la sua efficienza di eiezione. La relazione tra la gittata sistolica e il volume telediastolico è nota come legge di Frank-Starling:
- precarico, in quanto man mano che il cuore si riempie, le fibre si stirano e la massa di sangue rappresenta il carico applicato al muscolo prima che inizi la contrazione. Esiste un valore ottimale di precarico da cui il muscolo cardiaco è in grado di sviluppare la maggior forza contrattile;
- postcarico, rappresentato dalla pressione diastolica dell’aorta e delle arterie polmonari che il cuore deve vincere per potervi spingere il sangue.
Il cuore ha la capacità intrinseca di adattarsi a variazioni del volume di sangue venoso in ingresso: infatti, più aumenta il volume telediastolico, più le fibre si stirano e più aumentano la forza di contrazione e la gittata. Oltre un certo volume telediastolico però, la gittata diminuisce, indicando
quindi che il ventricolo è troppo dilatato ed è diminuita la sua efficienza di eiezione. La relazione tra la gittata sistolica e il volume telediastolico è nota come legge di Frank-Starling:
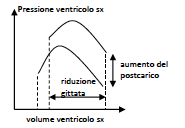
Mentre un aumento del precarico determina un aumento della gittata sistolica, un aumento del postcarico determina invece una riduzione della gittata sistolica:
Durante esercizio fisico, quando è richiesto un aumento della prestazione cardiaca, la noradrenalina rilasciata nel miocardio dal sistema simpatico determina uno spostamento di questa curva, con incremento dello stato inotropo e cronotropo:

Diagramma di Wiggers: descrive, rispetto all’asse del tempo, l’andamento dei vari parametri come l’ECG, le pressioni endocavitarie, i volumi di atri e ventricoli.
L’ECG consiste nella registrazione a livello cutaneo dell’attività elettrica globale del cuore, in funzione del tempo: la somma algebrica di tutti i p.a. generati nelle fibre miocardiche dà luogo infatti a specifiche fluttuazioni dotate di tensione elettrica, che possono essere registrate:
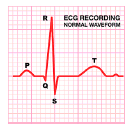
- complesso QRS: indica la depolarizzazione ventricolare ;
- onda T: indica la ripolarizzazione ventricolare;
- la frequenza cardiaca è ricavata dalla distanza tra due onde R successive;
- la morfologia del complesso QRS inoltre, è in grado di fornire info sul trofismo della muscolatura ventricolare o su eventuali asincronismi di trasmissione dell’impulso ai due ventricoli;
- possono essere individuati anche eventuali battiti ventricolari prematuri o eventuali “slivellamenti” nel tratto ST, che è indice di infarto del miocardio.
Metabolismo cardiaco e circolazione coronarica: il miocardio si contrae sfruttando l’energia liberata dalla scissione di ATP secondo la reazione ATP+H20=ADP+P+Energia.
L’ATP consumato deve quindi essere subito ricostituito: ciò avviene tramite la fosforilazione ossidativa nei mitocondri. Si ha consumo di O2 e si producono CO2 e acqua, liberando energia sottoforma di molecole di ATP. A differenza di quanto accade nel muscolo scheletrico, il miocardio non è in grado di utilizzare la glicolisi anaerobica: di conseguenza, il metabolismo miocardico dipende strettamente da una continua e adeguata fornitura di O2 da parte del circolo coronarico.
Quindi, un incremento delle richieste di O2 da parte del miocardio possono essere soddisfatte solo da un incremento della perfusione coronarica (iperemia metabolica).
Nell’uomo, a riposo, il flusso coronarico ammonta a 60-90ml per 100g di tessuto miocardico e nel corso dell’esercizio fisico può aumentare di 4-5 volte: questa capacità di adeguamento costituisce la “riserva coronarica”.
Il flusso coronarico è max durante la diastole e diminuisce fino a diventare negativo durante la sistole, a causa delle forze compressive esercitate dal cuore sui vasi durante la contrazione: questa situazione è però compensata da un’elevata densità capillare, che assicura un’estrazione massimale di O2.
Il consumo di O2 del cuore (MVO2), a riposo, è di 9ml/min per 100g di tessuto: la frazione necessaria al metabolismo basale miocardico è circa il 19%, quella necessaria all’attività elettrica è lo 0,05% mentre il restante 80% è suddiviso tra frequenza cardiaca, sviluppo di pressione parietale e livello di inotropismo.
L’ATP consumato deve quindi essere subito ricostituito: ciò avviene tramite la fosforilazione ossidativa nei mitocondri. Si ha consumo di O2 e si producono CO2 e acqua, liberando energia sottoforma di molecole di ATP. A differenza di quanto accade nel muscolo scheletrico, il miocardio non è in grado di utilizzare la glicolisi anaerobica: di conseguenza, il metabolismo miocardico dipende strettamente da una continua e adeguata fornitura di O2 da parte del circolo coronarico.
Quindi, un incremento delle richieste di O2 da parte del miocardio possono essere soddisfatte solo da un incremento della perfusione coronarica (iperemia metabolica).
Nell’uomo, a riposo, il flusso coronarico ammonta a 60-90ml per 100g di tessuto miocardico e nel corso dell’esercizio fisico può aumentare di 4-5 volte: questa capacità di adeguamento costituisce la “riserva coronarica”.
Il flusso coronarico è max durante la diastole e diminuisce fino a diventare negativo durante la sistole, a causa delle forze compressive esercitate dal cuore sui vasi durante la contrazione: questa situazione è però compensata da un’elevata densità capillare, che assicura un’estrazione massimale di O2.
Il consumo di O2 del cuore (MVO2), a riposo, è di 9ml/min per 100g di tessuto: la frazione necessaria al metabolismo basale miocardico è circa il 19%, quella necessaria all’attività elettrica è lo 0,05% mentre il restante 80% è suddiviso tra frequenza cardiaca, sviluppo di pressione parietale e livello di inotropismo.
Lavoro e rendimento cardiaco: nel corso della sistole di ogni battito, il cuore sviluppa un lavoro che è suddivisibile in due componenti per ciascun ventricolo:
- volumetrica (wv), calcolata tramite l’integrale della curva pressione-volume, oppure tramite il prodotto della pressione media per la gittata del ventricolo (nel ventr.sx: wv=0,9 J ; nel ventr.dx: wv=0,1 J);
- cinetica (wc), calcolata facendo: 0,5 ∙ massa di sangue ad ogni qc ∙ velocità del sangue in aorta e arteria polmonare. Per ciascun ventricolo, wc=0,014 J.
Il lavoro totale ad ogni battito sarà quindi dato dalla somma di wv+wc dei due ventricoli, e corrisponde a 1,176 J.
Conoscendo il dispendio energetico cardiaco (MVO2) e il lavoro svolto dal cuore, si ottiene il rendimento cardiaco (η): η=lavoro∙min/MVO2
Considerando una frequenza a riposo di 70 battiti/min, si ha η= 12,4%
- volumetrica (wv), calcolata tramite l’integrale della curva pressione-volume, oppure tramite il prodotto della pressione media per la gittata del ventricolo (nel ventr.sx: wv=0,9 J ; nel ventr.dx: wv=0,1 J);
- cinetica (wc), calcolata facendo: 0,5 ∙ massa di sangue ad ogni qc ∙ velocità del sangue in aorta e arteria polmonare. Per ciascun ventricolo, wc=0,014 J.
Il lavoro totale ad ogni battito sarà quindi dato dalla somma di wv+wc dei due ventricoli, e corrisponde a 1,176 J.
Conoscendo il dispendio energetico cardiaco (MVO2) e il lavoro svolto dal cuore, si ottiene il rendimento cardiaco (η): η=lavoro∙min/MVO2
Considerando una frequenza a riposo di 70 battiti/min, si ha η= 12,4%
Parametri per valutare la funzionalità cardiaca:
- la v max di incremento pressorio durante la contrazione ventricolare isovolumetrica;
- il “doppio prodotto”, ottenuto moltiplicando il valore sistolico della pressione arteriosa max per la frequenza cardiaca;
- la v max di incremento pressorio durante la contrazione ventricolare isovolumetrica;
- il “doppio prodotto”, ottenuto moltiplicando il valore sistolico della pressione arteriosa max per la frequenza cardiaca;
- l’indice “tensione-tempo”, calcolato sommando le aree sottese alla fase sistolica del tracciato delle pressioni endocavitarie del ventr.sx , registrato per 1min;
- frazione di eiezione, data dal rapporto percentuale tra volume di qc e volume telediastolico.
Di norma questo valore è compreso tra 55% e 80%: valori inferiori a 55% indicano una ridotta contrattilità miocardica.
- frazione di eiezione, data dal rapporto percentuale tra volume di qc e volume telediastolico.
Di norma questo valore è compreso tra 55% e 80%: valori inferiori a 55% indicano una ridotta contrattilità miocardica.
Gittata: è la quantità di flusso continuo di sangue che viene pompato dal cuore in un minuto, che varia in base alle richieste metaboliche dell’organismo.
La gittata (Q) =FC∙qc , dove FC=frequenza cardiaca e qc=gittata sistolica ventricolare.
In un individuo di media corporatura, a riposo, Q= 70battiti/min ∙ 75ml, ovvero 5litri/min.
In condizioni di lavoro, ad esempio durante esercizio fisico, si ha un aumento della FC e della qc: in
generale, la gittata cardiaca può aumentare di 3,5-5 volte rispetto alle condizioni di riposo, a causa dell’incremento di almeno 3 volte di FC e di almeno 2 volte di qc. In atleti olimpionici o in caso di esercizi massimali la gittata può arrivare anche a 35l/min.
La gittata (Q) =FC∙qc , dove FC=frequenza cardiaca e qc=gittata sistolica ventricolare.
In un individuo di media corporatura, a riposo, Q= 70battiti/min ∙ 75ml, ovvero 5litri/min.
In condizioni di lavoro, ad esempio durante esercizio fisico, si ha un aumento della FC e della qc: in
generale, la gittata cardiaca può aumentare di 3,5-5 volte rispetto alle condizioni di riposo, a causa dell’incremento di almeno 3 volte di FC e di almeno 2 volte di qc. In atleti olimpionici o in caso di esercizi massimali la gittata può arrivare anche a 35l/min.
Fattori esterni che influiscono su Q sono:
- l’aumento di postcarico, che porta a una diminuzione di Q;
- l’aumento di precarico, che porta ad un aumento di Q, secondo la legge di Frank-Starling.
Uno dei metodi per misurare la gittata cardiaca si basa sul principio di Fick, secondo cui la velocità di trasporto di un soluto da parte di un fluido è uguale al prodotto della velocità di flusso del fluido per la concentrazione di soluto nel fluido stesso. L’applicazione di questo principio consente di
misurare la gittata cardiaca se si conoscono il consumo di O2 misurato alla bocca (VO2) e il contenuto di O2 nel sangue arterioso (CaO2) e venoso misto (CvO2), quindi:
Q= VO2 / CaO2-CvO2.
- l’aumento di postcarico, che porta a una diminuzione di Q;
- l’aumento di precarico, che porta ad un aumento di Q, secondo la legge di Frank-Starling.
Uno dei metodi per misurare la gittata cardiaca si basa sul principio di Fick, secondo cui la velocità di trasporto di un soluto da parte di un fluido è uguale al prodotto della velocità di flusso del fluido per la concentrazione di soluto nel fluido stesso. L’applicazione di questo principio consente di
misurare la gittata cardiaca se si conoscono il consumo di O2 misurato alla bocca (VO2) e il contenuto di O2 nel sangue arterioso (CaO2) e venoso misto (CvO2), quindi:
Q= VO2 / CaO2-CvO2.
Continua a leggere:
- Successivo: Apparato respiratorio
- Precedente: Sistema cardiovascolare
Per approfondire questo argomento, consulta le Tesi:
- Employer Branding talenti ed alti potenziali: managerialità di nuova generazione
- Sensorialità autistica: una singolare esperienza del mondo. Comprendere il profilo di funzionamento sensoriale per progettare l'iter psicoeducativo
- Psicologia della comunicazione: rilevazioni quantitative di processi emotivi, attentivi, percettivi.
- La rieducazione del paziente cerebellare
- Il colore dei suoni. Un incantesimo percettivo.
Puoi scaricare gratuitamente questo appunto in versione integrale.
Forse potrebbe interessarti:
Sensorialità autistica: una singolare esperienza del mondo. Comprendere il profilo di funzionamento sensoriale per progettare l'iter psicoeducativo
 Ricevi informazioni sui nostri servizi, sulle offerte e non perdere news e consigli su università e lavoro.
Ricevi informazioni sui nostri servizi, sulle offerte e non perdere news e consigli su università e lavoro.
Login
Oppure utilizza il tuo account
 o
o
