
Gli Adenovirus
I secondi virus a circuito oro-fecale che prendiamo in considerazione sono gli Adenovirus, ed è il primo ciclo replicativo che esaminiamo di un DNA-virus.
Caratteristiche degli adenovirus
Fino ad ora abbiamo esaminato polio, rhino, tutti virus caratterizzati da avere come informazione genetica una molecola di RNA, questa è la prima volta che esaminiamo un virus con una doppia catena di DNA.
La maggior parte di questi virus sono trasmessi per via respiratoria, ma alcuni sierotipi sono trasmessi invece nel circuito oro-fecale e sono esattamente i sierotipi 40 e 41, ecco perché inseriamo gli Adeno a questo punto del corso.
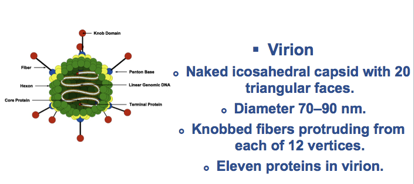
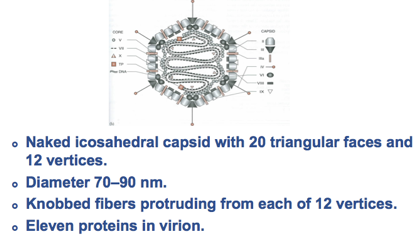
In questa prima diapositiva vediamo schematizzata una particella di Adenovirus che non ha rivestimento pericapsidico, sono infatti virus nudi e caratteristicamente ai vertici del capside icosaedrico passano dei prolungamenti che terminano con queste sferette, che vedete colorate in rosso, e vedremo che alla fine delle fibre è localizzato l’antirecettore, ossia il componente della particella virale che prenderà rapporto con il recettore presente nella cellula ospite.
Sono virus che esprimono la propria informazione genetica in maniera temporalmente molto ordinata. È un virus che promuove tutta una serie di eventi che sono correlati tra di loro in successione.
Da un unico promotore vengono trascritti quei metageni che portano alla sintesi delle proteine tardive. Forma quindi 5 messaggeri tardivi “LATE”.
Hanno un diametro di 60/90 nm, un icosaedro caratterizzato da 12 vertici ed ha un'informazione genetica di 30/36 Kbp, quindi un'informazione più grande rispetto ai virus che abbiamo fino ad oggi esaminato; riesce grazie a questa informazione a sintetizzare circa 50 proteine.
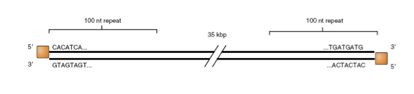
Qui vediamo analizzato il genoma formato da DNA bicatenario e la sua caratteristica è quella di avere una proteina (quadratino arancione) localizzata
agli estremi 5’ di entrambe le catene. Un'altra caratteristica che esamineremo guardando le modalità di replicazione del genoma degli adeno è che ogni catena presenta alle sue estremità, 3’ e 5’ mostra delle sequenze di un centinaio di desossiribonucleotidi, che sono caratterizzati da regioni inversamente ripetute e che possono tra loro appaiarsi legandosi per complementarietà.
La replicazione degli adenovirus
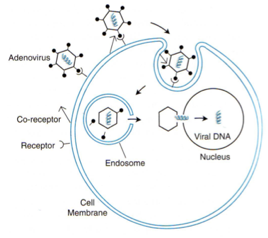
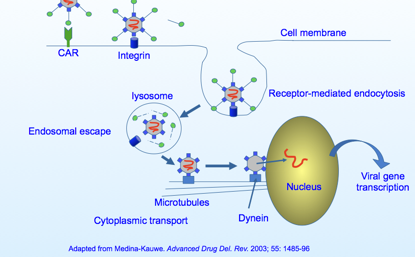
Qua vediamo illustrato in maniera schematica il ciclo replicativo di questi DNA-virus. Ecco qua una particella di Adeno, ci sono queste fibre che si proiettano ai vertici e terminano con un ingrossamento che prende rapporto col recettore della cellula ospite, ma un solo recettore non è sufficiente, infatti il virus per potersi legare alla cellula richiede l’intervento di un secondo recettore che funge da “corecettore”, quindi, quando avviene questo incontro con i 2 recettori il virus riesce a penetrare per endocitosi, l’acidificazione a livello del lisosoma libera il virus nel citoplasma, questo si dirige verso il nucleo e finalmente viene localizzato il genoma a DNA, che potrà così essere trascritto per arrivare alla sintesi delle proteine virali; queste proteine serviranno per formare nuovi genomi e sintetizzare le proteine strutturali. Questi elementi assemblandosi danno origine a nuove particelle di Adenovirus.Sono virus che esprimono la propria informazione genetica in maniera temporalmente molto ordinata. È un virus che promuove tutta una serie di eventi che sono correlati tra di loro in successione.
Partiamo dall'inizio di questo ciclo replicativo. Ecco qui la fibra che termina con questo rigonfiamento, dove vi ho detto essere localizzato l'antirecettore, e il recettore presente sulla cellula sono di solito cellule epiteliali (nel caso dei sierotipi 40e 41) dove è presente questa proteina “CAR” (in rosso); noi non sappiamo ancora quale è il ruolo fisiologico di questa proteina, ed è chiamata così perché è un recettore sia per i Coxsackievirus sia per gli adenovirus, una volta che è avvenuto questo legame con il CAR è necessario un secondo recettore (barrette nere che terminano con un rigonfiamento), si legano alla base del pentone e così stabilizzano ulteriormente il legame del virus con la cellula ospite e consentono l’inizio dell’endocitosi.
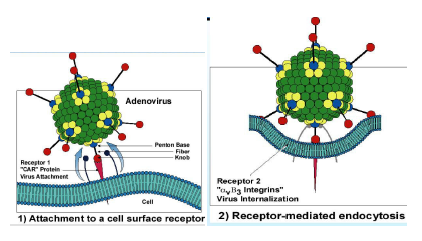
Quindi un legame con CAR, poi con un secondo corecettore il quale è un'integrina presente sulla cellula ospite, questa si lega col corecettore che si trova alla base della fibra; a questo punto avvenuto questo doppio legame, la particella virale viene adsorbita si muove sulla membrana citoplasmatica finché incontra una particolare porzione della membrana dove si può introflettere, qui si forma una vescicola di endocitosi. Questa vescicola sta in ambiente acido, il che consente la liberazione delle particelle virali nel citoplasma. Una volta arrivata, grazie all’aiuto di una proteina cellulare chiamata “Dineina” si muove lungo i microtubuli per arrivare finalmente nel nucleo, dove viene liberato il genoma virale che potrà dare avvio, sfruttando la RNA-pol DNA dipendente dell'ospite, ai processi che formeranno i messaggeri virali.
Cosa comporta questo processo di trascrizione?
Abbiamo parlato di un'informazione abbondante di DNA che porta alla formazione di circa 50 proteine.
Vediamo che tutto il genoma degli adeno viene sfruttato e trascritto per arrivare alla sintesi di metageni, vengono trascritte regioni complementari dello stesso tratto di DNA e questo porterà alla formazione di m-RNA tra loro complementari. Vedremo che questo sarà un punto di partenza per la stimolazione dell'Interferon; per quanto riguarda i promotori riconosciuti dalla RNA-pol 2 cellulare, responsabile quindi della formazione dei trascritti virali, ci sono dei promotori “precoci” , questi portano alla sintesi di questi metageni: E1A –E1B E2A-E2B-E3-E4 , ci sono poi anche altri 2 metageni che chiamiamo “precoci-ritardati” il messaggero 4A2- il 9 (scritti in numeri romani). E abbiamo sistemato i promotori che portano alla sintesi dei messaggeri precoci e precoci ritardati. Da un unico promotore vengono trascritti quei metageni che portano alla sintesi delle proteine tardive. Forma quindi 5 messaggeri tardivi “LATE”.
Quale è la caratteristica saliente della replicazione degli adenovirus? Una volta che da questi promotori si sono formati dei messaggeri, questi sottostanno ad un numero veramente elevato di splicing (fenomeno scoperto appunto negli adeno) e vedremo come questo splicing svolge un ruolo determinante per la sintesi di tutte le proteine necessarie per il suo ciclo replicativo. Ci sono altri 2 m-RNA “VA” che anziché essere trascritti dalla polimerasi di tipo 2 sono trascritti da quella di tipo 3, e trascrivono per proteine che sono molto importanti per gli attacchi del virus al sistema immuno-competente.
Ho fatto una sorta di introduzione alla diapositiva successiva:

ci mostra su quali catene vengono sintetizzati gli m-RNA degli adenovirus.
Vi parlavo prima di messaggeri precoci come E1A –E1b che sono localizzati da sinistra verso destra (si muovono in direzione 5'-3'). Di questi messaggeri precoci E1A-E1B-E3 vengono sintetizzati nella catena destra, mentre i trascritti E2A-E2B-E4 sono trascritti sulla catena sinistra. Abbiamo in tutto 4 messaggeri.
I promotori precoci tardivi sono la proteina 4A2 che viene trascritta nella catena sinistra mentre l'altra proteina, 9, viene trascritta nella destra. A partire da un unico promotore, si ha l'avvio per la sintesi di tutti i messaggeri LATE che saranno poi proteine strutturali e sono caratterizzate da avere l'inizio tutte in questa stessa regione chiamata “leader”.
I promotori precoci tardivi sono la proteina 4A2 che viene trascritta nella catena sinistra mentre l'altra proteina, 9, viene trascritta nella destra. A partire da un unico promotore, si ha l'avvio per la sintesi di tutti i messaggeri LATE che saranno poi proteine strutturali e sono caratterizzate da avere l'inizio tutte in questa stessa regione chiamata “leader”.
Quale è la proteina fondamentale che permette l'espressione degli adenovirus?
Questa proteina è la E1A; perché?
È l'unica che può essere trascritta solo da fattori di trascrizione cellulari, una volta che questo DNA entra nel nucleo della cellula, solo E1A può essere trascritto usando fattori cellulari e quindi di conseguenza quando questo DNA arriva nel nucleo avremo soltanto la formazione della proteina E1A. Se questa proteina non viene tradotta il virus non può esprimersi. Perché è cosi importante il suo ruolo?
Il virus va ad infettare delle cellule epiteliali, queste cellule sono sia cellule differenziate sia cellule quiescenti, cosa comporta questo? Il virus entra va a replicare nel nucleo ed ha estremo bisogno di collaborazione della cellula ospite per riuscire a replicare il proprio DNA. Una cellula quiescente non è pronta a replicare il DNA, è quindi in una fase di arresto; il virus trova quindi un ambiente cellulare che non corrisponde a quelle che sono le sue esigenze, e quale è il ruolo di E1A? È quello di dare un calcio alla cellula e dirgli di replicarsi per avere tutto quel materiale utile per la sua duplicazione. Quindi se non di esprime E1A la cellula non entra in fase S.
Gli adenovirus quindi necessitano di certo quantitativo di proteine dell'ospite e devono e spingere la cellula a produrre un certo numero di precursori ad esempio nucleotidi che il virus non è capace di sintetizzare da solo; tutto questo è realizzato dalla proteina E1A.
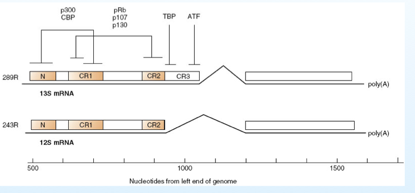
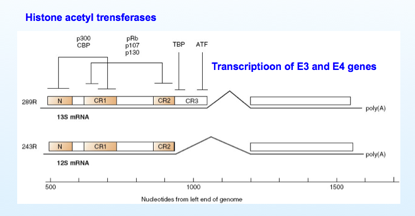
Come vedete nell'immagine da questo promotore, in seguito a splicing, si originano tutta una serie di 5 messaggeri (trascritti),e di questi hanno un ruolo determinante “289R”e “243R” (R sta per residui Aa) e sono conosciuti nel loro funzionamento; in seguito allo splicing la proteina “289R” risulta più lunga rispetto alla proteina “243R”. Vediamo quali domini possiamo riconoscere in questa proteina E1A, che come ripeto ha il compito di far entrare la cellula in fase S. In entrambe le proteine troviamo le sequenze “CR1”e “CR2” (CR=regione conservata), poi abbiamo una sequenza N ammino-terminale e la proteina più grande “289R” oltre ad avere CR1-CR2 ed N possiede anche la sequenza CR3.
Con chi reagiscono queste proteine tramite i segmenti CR1 e CR2? Con questi segmenti, queste proteine sono in grado di reagire con la famiglia delle proteine pRb –p107 e p130 (retinoblastoma). Queste proteine Rb grazie alle porzioni CR1 e CR2 possono essere legate alla proteina E1A. Cosa succede? E perché è importante avviare la cellula in fase S? Quando una cellula è ferma la fase S è repressa proprio da una proteina Rb che lega il fattore E2F, fattore di trascrizione, che permette di trascrivere tutti quei geni che portano la cellula ad entrare in S, quindi dei determinanti importanti per promuovere la duplicazione del DNA.
In un ciclo cellulare normale Rb si lega al fattore E2F e tiene la cellula ferma in G0, l'ingresso in fase S è bloccato. Stimoli, fattori di crescita, inducono la cellula a riprendere il ciclo replicativo; ecco che una serie di trasmissioni del segnale portano ad attivare una ciclino-chinasi dipendente che ha il compito di fosforilare Rb, vedete qui il passaggio, e una volta che Rb è stata fosforilata E2F è libero di funzionare e quindi di attivare la trascrizione di quei geni che portano la cellula alla fase S.
Con chi reagiscono queste proteine tramite i segmenti CR1 e CR2? Con questi segmenti, queste proteine sono in grado di reagire con la famiglia delle proteine pRb –p107 e p130 (retinoblastoma). Queste proteine Rb grazie alle porzioni CR1 e CR2 possono essere legate alla proteina E1A. Cosa succede? E perché è importante avviare la cellula in fase S? Quando una cellula è ferma la fase S è repressa proprio da una proteina Rb che lega il fattore E2F, fattore di trascrizione, che permette di trascrivere tutti quei geni che portano la cellula ad entrare in S, quindi dei determinanti importanti per promuovere la duplicazione del DNA.
In un ciclo cellulare normale Rb si lega al fattore E2F e tiene la cellula ferma in G0, l'ingresso in fase S è bloccato. Stimoli, fattori di crescita, inducono la cellula a riprendere il ciclo replicativo; ecco che una serie di trasmissioni del segnale portano ad attivare una ciclino-chinasi dipendente che ha il compito di fosforilare Rb, vedete qui il passaggio, e una volta che Rb è stata fosforilata E2F è libero di funzionare e quindi di attivare la trascrizione di quei geni che portano la cellula alla fase S.
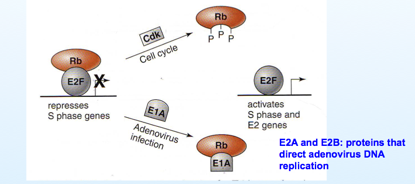
Quindi Rb è una proteina determinante nel regolare il ciclo cellulare. Cosa c'entra con gli adeno? Il virus entra, e deve necessariamente trascrivere un messaggero, questo messaggero porta alla sintesi di E1A che ha il compito di legarsi alla proteina Rb (sostituisce la Cdk), la sequestra e il risultato è analogo a quello che abbiamo appena esaminato, E2F è libero e tutta una serie di fattori di trascrizione vengono liberati, la cellula può entrare in S e prepararsi alla replicazione del proprio DNA. Inoltre questi fattori di trascrizione sono responsabili della trascrizione di altre 2 proteine virali, che sono la E2A- E2B, che vedremo essere responsabili della formazione di nuove molecole di DNA virale.
A seguito del funzionamento dei fattori E2F, la cellula decide di prendere la via apoptotica, però è un rischio, infatti se anche la cellula muore, anche la replicazione del virus sarà inattivata e allora bisogna correre ai ripari… in che modo? Arginando, stoppando la via apoptotica. Abbiamo esaminato questo aspetto in cui il legame con Rb induce la cellula ad entrare in fase S; ma l'attività della proteina E1A non si limita a questo, infatti con la sua porzione CR3 (presente solo nella proteina 289R) è in grado di attivare altri trascritti virali che portano alle proteine E3 E4, mentre con la porzione CR1 e con la porzione N è in grado di legare delle proteine cellulari, p300 e CBP che sono delle acetiltransferasi. Cosa fanno alle proteine attivate dall'E1A? Sappiamo che gli istoni hanno il compito di staccare i promotori sul DNA e questo disturba la loro trascrizione, ostacola i legami dei fattori di attivazione della trascrizione al promotore, ecco che grazie al legame tra la proteina E1A a queste attività enzimatiche, le attiva, e il risultato è che gli istoni vengono acetilati, e così assumono una conformazione molto più lassa, che facilita il riconoscimento, o meglio la localizzazione sui promotori dei fattori di attivazione della trascrizione. Tutto quanto deve essere trascritto all' inizio della fase S, quindi abbiamo da una parte l'intervento su Rb, che dice alla cellula puoi entrare in S, e come può facilitare ulteriormente la formazione di trascritti che avvengono nella cellula all'inizio della fase S? Rende più lassi gli istoni grazie all'attivazione di proteine come CBP e p300, che acetilano gli istoni e rendono più facile la trascrizione.
A seguito del funzionamento dei fattori E2F, la cellula decide di prendere la via apoptotica, però è un rischio, infatti se anche la cellula muore, anche la replicazione del virus sarà inattivata e allora bisogna correre ai ripari… in che modo? Arginando, stoppando la via apoptotica. Abbiamo esaminato questo aspetto in cui il legame con Rb induce la cellula ad entrare in fase S; ma l'attività della proteina E1A non si limita a questo, infatti con la sua porzione CR3 (presente solo nella proteina 289R) è in grado di attivare altri trascritti virali che portano alle proteine E3 E4, mentre con la porzione CR1 e con la porzione N è in grado di legare delle proteine cellulari, p300 e CBP che sono delle acetiltransferasi. Cosa fanno alle proteine attivate dall'E1A? Sappiamo che gli istoni hanno il compito di staccare i promotori sul DNA e questo disturba la loro trascrizione, ostacola i legami dei fattori di attivazione della trascrizione al promotore, ecco che grazie al legame tra la proteina E1A a queste attività enzimatiche, le attiva, e il risultato è che gli istoni vengono acetilati, e così assumono una conformazione molto più lassa, che facilita il riconoscimento, o meglio la localizzazione sui promotori dei fattori di attivazione della trascrizione. Tutto quanto deve essere trascritto all' inizio della fase S, quindi abbiamo da una parte l'intervento su Rb, che dice alla cellula puoi entrare in S, e come può facilitare ulteriormente la formazione di trascritti che avvengono nella cellula all'inizio della fase S? Rende più lassi gli istoni grazie all'attivazione di proteine come CBP e p300, che acetilano gli istoni e rendono più facile la trascrizione.
Vi stavo dicendo anche che E1A programma la cellula perché sia adatta al virus, però creava anche un problema; induce la cellula ad una via apoptotica.
Perché succede questo? Vediamolo nei dettagli:
• Questo succede perché la proteina E1A non fa volutamente tale processo, ma attiva vari trascritti che portano anche all'attivazione della proteina p53.
• Questo succede perché la proteina E1A non fa volutamente tale processo, ma attiva vari trascritti che portano anche all'attivazione della proteina p53.
Quale è il compito di questa proteina? È quello di controllare il ciclo replicativo, infatti può sia attivare che reprimere processi di trascrizione.
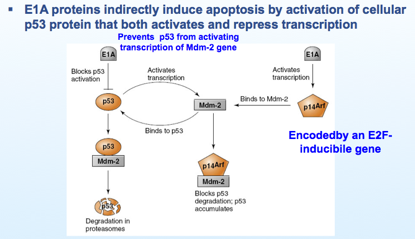
Come vanno le cose normalmente? Questa è la p53 che se attivata e se si accumula dice alla cellula prendi la via apoptotica. Una cellula normale va incontro a danneggiamento del DNA e ci sono tutta una serie di segnali accumulo e attivazione della p53 e apre la porta all'apoptosi. Come viene regolata normalmente la p53? Viene regolata da un’altra proteina chiamata MDM2. Infatti la p53 attiva la MDM2 che a sua volta si lega alla p53 e ne determina la degradazione nei proteasomi. In poche parole la MDM2 controlla la concentrazione di p53. A sua volta da chi è regolata la MDM2? È regolata da un'altra proteina che è la p14arf che si lega alla MDM2 , e in questa maniera MDM2 viene sequestrata e si avrà l’accumulo di p53.
Cosa succede quando la proteina E1A virale degli adeno libera il fattore E2F ?
Ricordate che E1A deve liberare questo fattore per indurre la cellula ad entrare in fase S e succede che induce anche la trascrizione della proteina p14, più proteina p14 viene prodotta, maggiore sarà il suo legame alla MDM2, e di conseguenza avremo accumulo della p53.
Quindi abbiamo visto che a causa della liberazione dei fattori di trascrizione viene anche indotto il gene della p53, quindi maggior sequestro della proteina MDM2, che ha come ultima conseguenza l'aumento della p53 che non viene degradata. Ma l’intervento di E1A non si limita a questo. E1A è capace di bloccare l’attivazione da parte di p53 della proteina MDM2, bloccando questa attivazione, il risultato sarà sempre quello di diminuire la concentrazione di MDM2 con conseguente accumulo di p53 e quindi segnale per la cellula di imboccare la via apoptotica. Agisce sia indirettamente che direttamente.
Come vanno le cose normalmente? Questa è la p53 che se attivata e se si accumula dice alla cellula prendi la via apoptotica. Una cellula normale va incontro a danneggiamento del DNA e ci sono tutta una serie di segnali accumulo e attivazione della p53 e apre la porta all'apoptosi. Come viene regolata normalmente la p53? Viene regolata da un’altra proteina chiamata MDM2. Infatti la p53 attiva la MDM2 che a sua volta si lega alla p53 e ne determina la degradazione nei proteasomi. In poche parole la MDM2 controlla la concentrazione di p53. A sua volta da chi è regolata la MDM2? È regolata da un'altra proteina che è la p14arf che si lega alla MDM2 , e in questa maniera MDM2 viene sequestrata e si avrà l’accumulo di p53.
Cosa succede quando la proteina E1A virale degli adeno libera il fattore E2F ?
Ricordate che E1A deve liberare questo fattore per indurre la cellula ad entrare in fase S e succede che induce anche la trascrizione della proteina p14, più proteina p14 viene prodotta, maggiore sarà il suo legame alla MDM2, e di conseguenza avremo accumulo della p53.
Quindi abbiamo visto che a causa della liberazione dei fattori di trascrizione viene anche indotto il gene della p53, quindi maggior sequestro della proteina MDM2, che ha come ultima conseguenza l'aumento della p53 che non viene degradata. Ma l’intervento di E1A non si limita a questo. E1A è capace di bloccare l’attivazione da parte di p53 della proteina MDM2, bloccando questa attivazione, il risultato sarà sempre quello di diminuire la concentrazione di MDM2 con conseguente accumulo di p53 e quindi segnale per la cellula di imboccare la via apoptotica. Agisce sia indirettamente che direttamente.
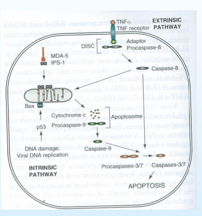
Cosa comporta l'accumulo di p53? Si ha la formazione di oligomeri formati da proteine Bax, dopo che abbiamo avuto tutta una serie di trasmissioni di segnali, ecco che Bax si lega alla membrana dei mitocondri, si formano dei fori nella struttura e liberano il citocromo C, che andrà ad attivare una procaspasi 9 e a questo punto innesca una reazione a catena fino ad attivare le caspasi 3 e 7 e la cellula andrà incontro al processo apoptotico e finirà in pezzettini.
Per quando riguarda gli adeno, siamo arrivati ad un effetto involontario della E1A che porta per una via diretta e una indiretta ad accumulo di p53, facendosi mancare così il tempo per compiere il ciclo replicativo, perché la cellula muore.
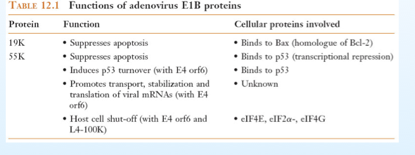
Ecco però che una seconda proteina entra in gioco ad aiutare il virus, la E1B e blocca la via apoptotica che involontariamente la E1A ha attivato; in questo modo il virus potrà mandare avanti il suo ciclo replicativo. Ancora una volta attraverso lo splicing, E1B viene divisa in più proteine e le più famose sono: 19K e 55K. Come fanno queste proteine a bloccare l’apoptosi? La 19K la sopprime in questa maniera: vi ho detto prima che Bax è un oligomero formato da tante proteine che si lega alla membrana del mitocondrio, il ruolo di E1B è quello di impedire la formazione di questi oligomeri, si lega a Bax e blocca la via apoptotica. L’altra proteina, la 55k, si lega alla p53 e ne reprime la trascrizione, quindi blocca la sua produzione. Ma non si limita a fare questo, ed insieme ad un’altra proteina virale la E4orf6 , fa si che la p53 venga velocemente degradata a livello dei proteasomi. Quindi questi 2 interventi diminuiscono drasticamente i livelli di p53, dall’altra parte la 19K ha evitato il contatto con Bax e la membrana del mitocondrio, quindi la cellula evita la via apoptotica e il virus realizza il suo intento.
Ecco però che una seconda proteina entra in gioco ad aiutare il virus, la E1B e blocca la via apoptotica che involontariamente la E1A ha attivato; in questo modo il virus potrà mandare avanti il suo ciclo replicativo. Ancora una volta attraverso lo splicing, E1B viene divisa in più proteine e le più famose sono: 19K e 55K. Come fanno queste proteine a bloccare l’apoptosi? La 19K la sopprime in questa maniera: vi ho detto prima che Bax è un oligomero formato da tante proteine che si lega alla membrana del mitocondrio, il ruolo di E1B è quello di impedire la formazione di questi oligomeri, si lega a Bax e blocca la via apoptotica. L’altra proteina, la 55k, si lega alla p53 e ne reprime la trascrizione, quindi blocca la sua produzione. Ma non si limita a fare questo, ed insieme ad un’altra proteina virale la E4orf6 , fa si che la p53 venga velocemente degradata a livello dei proteasomi. Quindi questi 2 interventi diminuiscono drasticamente i livelli di p53, dall’altra parte la 19K ha evitato il contatto con Bax e la membrana del mitocondrio, quindi la cellula evita la via apoptotica e il virus realizza il suo intento.
Adesso vedremo delle proteine implicate nella duplicazione del genoma virale: una volta che E1A ha indotto la cellula ad entrare in S, vengono trascritte le prime proteine virali e può iniziare la duplicazione del DNA. Le proteine fondamentali implicate in questo processo sono la E2A ed E2B che danno origine a 3 proteine che svolgono un ruolo predominante nella duplicazione virale degli adeno. La proteina E2A darà origine ad una proteina che si lega al DNA a singola catena, vedremo perché questo è indispensabile; i prodotti di E2B sono una DNA polimerasi (non usa quella cellulare), ed una proteina pre-terminale.
Voi sapete che in un normale processo di duplicazione del DNA, in una delle catene la sintesi è continua e nell'altra catena è discontinua, con formazione di primer che poi verranno rimossi ecc.; gli adeno hanno scelto una strategia di replicazione del proprio DNA particolare, qui la vediamo schematizzata:
Voi sapete che in un normale processo di duplicazione del DNA, in una delle catene la sintesi è continua e nell'altra catena è discontinua, con formazione di primer che poi verranno rimossi ecc.; gli adeno hanno scelto una strategia di replicazione del proprio DNA particolare, qui la vediamo schematizzata:
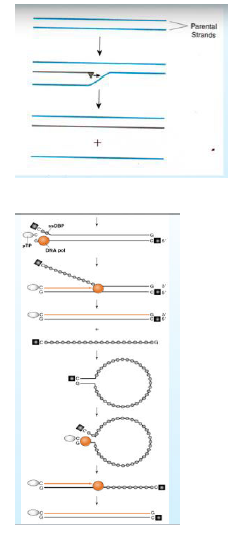
vediamo la DNA polimerasi che scorre e vedete come copia una catena e man mano che copia (quella in nero è la catena nuova) discosta la catena parentale; quale sarà il risultato, la produzione di una molecola di DNA di nuova sintesi con una catena stampo e una neo sintetizzata, mentre è stata dislocata una delle due catene parentali.
Vediamolo nei dettagli: cosa rende possibile la via della duplicazione del genoma? È necessaria la sintesi di una DNA polimerasi virus dipendente, virus-specifica, e di una proteina (sferetta bianca) chiamata proteina pre-terminale; che funzione ha? Ha il compito di fungere da primer, presenta un residuo di Serina, che espone l'OH (in precedenza questa proteina reagisce con un Deossi CTP, si lega con legame fosfodiesterico e viene degradato a CMP ) questo CMP sarà il punto di partenza della sintesi del DNA virale (CMP si accoppia con la G dello stampo che trova sul DNA). A questo punto la polimerasi inizia a sintetizzare una delle due catene di DNA e mentre fa questo discosta l’altra, man mano che la discosta la riveste della proteina che si lega al DNA a singola catena.
Ripartiamo dall'inizio: abbiamo una molecola bicatenaria di DNA virale, ad una estremità si lega la DNA polimerasi, neo sintetizzata,questa polimerasi utilizza un innesco proteico (proteina pre-terminale), questa espone un OH libero per formare un legame fosfodiesterico, segue l’aggancio della polimerasi che può iniziare a scorrere in maniera continua una delle due catene parentali, l’altra catena viene allontana e rivestita della proteina pre-terminale.
Quale è il risultato? La formazione di una catena di DNA neo sintetizzata (quella in arancione e quella scura è la parentale) e una catena parentale che è stata allontanata e rivestita dalla proteina pre-terminale specifica; però il virus ha copiato solo una catena e questo non è molto conveniente perché ha fretta di fare nuovi genomi da utilizzare per sintetizzare nuove particelle virali, quindi cosa succede? Il DNA degli adeno è caratterizzato dall'avere delle sequenze inversamente ripetute alle estremità, queste consentono a questa molecola di DNA di chiudersi agli estremi, si forma questa struttura che può ricordare una “padella con una manico”, perché è importante formare questa struttura? Questa zona forma una doppia catena che ricorda la porzione terminale del genoma virale, si ricrea una doppia catena di DNA che sarà riconosciuta dalla polimerasi, dalla proteina pre-terminale e si ripete quello che abbiamo visto all’inizio della sintesi del DNA; ed ecco che la polimerasi incontra un DNA bicatenario lo riconosce e comincia a copiare questo primo tratto bicatenario e il risultato che anche questa catena inizialmente spostata, viene copiata. Si sono formati due genomi distinti.
Cosa succede alla proteina innesco? Durante le fasi di maturazione del virus questa proteina pre-terminale viene clivata in una proteina più piccola chiamata proteina terminale che è legata all'estremità 5'di tutte e 2 le catene del genoma degli adeno.
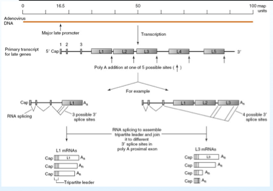
Adesso dobbiamo sintetizzare le proteine virali. Tutte le proteine tardive derivano tutte da uno stesso promotore, che darà origine a 5 messaggeri L1 L2 L3 L4 L5 , cosa determina la formazione di questi messaggeri? Il trascritto ha la possibilità di fermarsi in 5 differenti siti di poliadenilazione, e attraverso lo splicing saranno circa una ventina gli m-RNA che daranno origine alle proteine strutturali; come viene organizzato questo splicing? Qua abbiamo un esempio per il segmento L1 ed L3. Questo è L1.
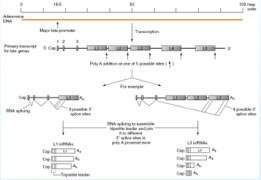
Il sito di poliadenilazione è tra L1 ed L2 e sono possibili 3 differenti siti di splicing, quindi da L1 possono essere prodotti 3 diversi m-RNA con sintesi di proteine diverse. La stessa cosa avviene in L3, anche qui rimozione degli introni, e i 3 esoni porteranno alla formazione di un leader, a livello del segmento L3 ci sono 4 siti di splicing, di conseguenza saranno prodotti i messaggeri per la sintesi di 4 diverse proteine. Lo splicing amplifica la possibilità di produrre proteine.
Perché queste sintesi avvengono dopo la duplicazione del DNA? Vi ho detto che il virus è molto preciso quindi non spreca nulla. Nella fase iniziale il promotore per queste proteine è estremamente debole e non viene praticamente riconosciuto, appena avviene la duplicazione, il DNA neo sintetizzato interagisce con delle proteine che alterano il DNA (causano dei cambiamenti di conformazione), modificano il promotore e lo rendono forte.
Abbiamo inoltre un'altra proteina la IV (quarta) che legandosi al promotore, incrementa l’attività della RNA polimerasi RNA dipendente. Ci sono poi 2 proteine virali E1B-55K e la E4orf6. Queste vanno nel nucleo riconoscono i messaggeri che hanno un leader in 5'e ne favoriscono il trasferimento nel citoplasma, quindi sono responsabili del trasporto selettivo dei messaggeri late caratterizzati dalla presenza di una sequenza leader all'estremità 5’.
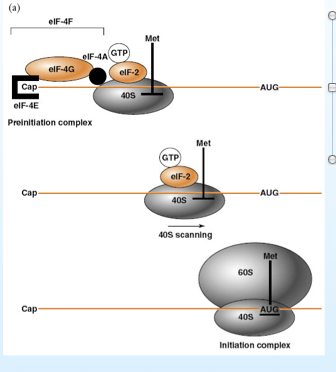
Non finisce qui l’importanza di questo leader. Infatti una volta arrivati nel citoplasma saranno in competizione coi messaggeri cellulari, come vince la battaglia? Un ruolo fondamentale nella sintesi normale è svolto da fattori come eif -4f, abbiamo il 4e che riconosce il cappuccio, il fattore 4a e il 4g, concorrono tutti alla formazione del complesso iniziale della sintesi proteica, è richiesto un f-met, una subunità ribosomiale 40s , e una volta che si assembla il ribosoma può scorrere in uno stampo fino ad incontrare il primo AUG ed iniziare la sintesi.
Non finisce qui l’importanza di questo leader. Infatti una volta arrivati nel citoplasma saranno in competizione coi messaggeri cellulari, come vince la battaglia? Un ruolo fondamentale nella sintesi normale è svolto da fattori come eif -4f, abbiamo il 4e che riconosce il cappuccio, il fattore 4a e il 4g, concorrono tutti alla formazione del complesso iniziale della sintesi proteica, è richiesto un f-met, una subunità ribosomiale 40s , e una volta che si assembla il ribosoma può scorrere in uno stampo fino ad incontrare il primo AUG ed iniziare la sintesi.
Cosa fa il virus? Una serie di proteine degli adeno sono in grado di defosforilare il fattore 4f; una volta che è defosforilato i messaggeri cellulari perdono la possibilità di essere tradotti. Grazie all'avere un leader in 5’, i messaggeri virali riescono a superare questo blocco tra questo leader e la porzione18 s del ribosoma c’è infatti complementarietà, il leader si lega alla sub 40s e la sintesi proteica con stampo messaggeri virali riesce a procedere. I messaggeri cellulari sono bloccati e quelli virali liberi.
Ricordiamo che nel mentre il virus ha bloccato l'apoptosi, e come fa ad uscire dalla cellula? Lo fa sintetizzando una proteina di morte. Fino ad ora ha sintetizzato le sue proteine strutturali, sono pronti i genomi per essere impaccati, però perché il virus possa arrivare ad infettare altre cellule deve essere liberato. La proteina di cui parlavamo prima induce la cellula ad entrare in apoptosi, ma non si sa con quale modalità avvenga questa induzione da parte della proteina, cmq il risultato finale è la rottura della cellula e la diffusione nell'ambiente extracellulare.
La risposta immunitaria agli adenovirus
Gli adeno sono diffusi per via respiratoria, ma alcuni sierotipi sono capaci di trasmissione nel circuito oro-fecale. La duplicazione degli adeno è lenta (48 h) che conseguenze potete trarre? È 1 virus che si deve armare di molte difese per contrastare quelle dell'ospite.
Queste armi sono tante ed efficaci ma non inarrestabili. Il virus non vuole portare a morte l'ospite. Prima di tutto abbiamo la difesa contro l'interferon. Vi dicevo all'inizio che porzioni del DNA complementare stimolano la sintesi dell'interferon, quando la cellula viene avvisata da questa molecola, sintetizza una proteina chinasi RNA dipendente (PKR) che viene attivata. Come contrasta l'azione della PKR? Con una proteina trascritta dall'RNA-pol III che ha il compito di legarsi alla PKR, inattivandola; se questa viene inattivata il fattore EIF 2 potrà seguire il proprio ciclo che porta alla trifosforilazione del GTP e la sintesi proteica non viene alterata. Quindi il ruolo della proteina che chiamiamo VA è quello di legarsi alla PKR inattivare questa chinasi e rendere possibile il normale procedere delle sintesi proteiche delle cellule virus infette.
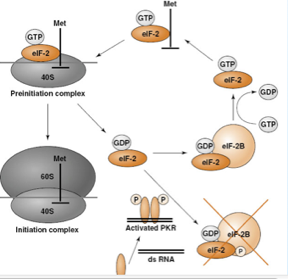
Queste armi sono tante ed efficaci ma non inarrestabili. Il virus non vuole portare a morte l'ospite. Prima di tutto abbiamo la difesa contro l'interferon. Vi dicevo all'inizio che porzioni del DNA complementare stimolano la sintesi dell'interferon, quando la cellula viene avvisata da questa molecola, sintetizza una proteina chinasi RNA dipendente (PKR) che viene attivata. Come contrasta l'azione della PKR? Con una proteina trascritta dall'RNA-pol III che ha il compito di legarsi alla PKR, inattivandola; se questa viene inattivata il fattore EIF 2 potrà seguire il proprio ciclo che porta alla trifosforilazione del GTP e la sintesi proteica non viene alterata. Quindi il ruolo della proteina che chiamiamo VA è quello di legarsi alla PKR inattivare questa chinasi e rendere possibile il normale procedere delle sintesi proteiche delle cellule virus infette.
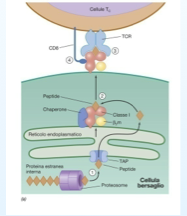
Un'altra difesa che abbiamo già nominato è l'attivazione della proteina della morte, che induce la cellula ad una via apoptotica. Un'altra difesa è quella di alcune proteine di interagire con le proteine MHC di classe I. Le proteine virali a livello del proteasoma vengono degradate vengono trasferite nel R.E. dove vengono legate alla proteina MHC I. A questo punto il complesso MHC I con l'antigene e l'aiuto di un chaperon viene esposto sulla superficie della cellula che richiama le proprie difese. Come fa a fermare questo? Gli adeno hanno evoluto un meccanismo che non permette alle proteine virali di essere riconosciute come estranee. Infatti una proteina virale, la E319K, ancorata al reticolo endoplasmatico, cattura le molecole MHC I le immobilizza e queste non possono migrare sulla superficie della cellula infetta.
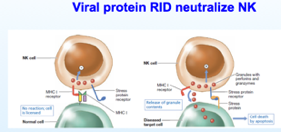
La furbizia del virus non finisce qui, ci sono altre difese da contrastare oltre ai linfociti T citotossici (immunità adattativa), infatti dobbiamo contrastare anche le Natural Killer (immunità innata) queste cellule controllano con una porzione (quella rossa) che ci sia una normale concentrazione di proteine MHC, un altro braccio (quello azzurro) se non riconosce nulla di strano non prende contatto con la cellula bersaglio (riconosce proteine da stress che evocano perforine e granzimi che distruggono la cellula infetta).
Ma cosa succede se incontra una cellula virus infetta? Molti virus sono in grado come gli adeno di non mostrare le proteine MHC sulla superficie; rimangono catturate al livello del R.E., quindi questa cellula avendo poche MHC passa il check-in. Inoltre non è ancora chiaro il meccanismo, ma si ritiene che una delle proteine della famiglia E3, chiamata RID, è in grado di internalizzare le proteine da stress che sono sulla superficie della cellula virus infetta, le porta all'interno della cellula in modo tale che il braccio azzurro della NT non riconosca neanche questi elementi e il virus possa permeare.
Continua a leggere:
- Successivo: I Rotavirus
- Precedente: I virus a trasmissione orale: l'epatite A
Dettagli appunto:
-
Autore:
Simone Pisu
[Visita la sua tesi: "Caratterizzazione molecolare ed eterogenicità delle varianti emoglobiniche in Sardegna"]
- Università: Università degli Studi di Cagliari
- Facoltà: Biologia
- Corso: Biologia Cellulare e Molecolare
- Esame: Virologia molecolare
- Docente: Prof. Marongiu
Altri appunti correlati:
- Microbiologia clinica
- Biologia applicata
- Microbiologia
- Microbiologia Medica
- Filologia della letteratura italiana
Per approfondire questo argomento, consulta le Tesi:
- Piante medicinali come ausilio per contrastare le patologie virali: tre casi-studio
- Contributo allo studio ed alla terapia dell'epatite C nel paziente HIV-positivo
- Memetica e nuovi media
- Peptidi overlapping derivanti da sequenze gH-omologhe di HSV-1 come inibitori dell'attività virale.
- Selezione di Mutanti HIV-1 Resistenti e Basi Molecolari della Resistenza di Nuovi Inibitori Non-Nucleosidici della Trascrittasi Inversa
Puoi scaricare gratuitamente questo appunto in versione integrale.
 Ricevi informazioni sui nostri servizi, sulle offerte e non perdere news e consigli su università e lavoro.
Ricevi informazioni sui nostri servizi, sulle offerte e non perdere news e consigli su università e lavoro.