
Il virus del morbillo
Il terzo virus, trasmesso per via respiratoria, che esaminiamo è il virus del morbillo, una malattia che normalmente riguarda l’infanzia e il virus che la causa è un paramixovirus.
Com’è fatta una particella di paramixovirus
Guardando questa figura immediatamente ci si rende conto che è un virus provvisto di envelope, il quale è costituito da glicoproteine e precisamente abbiamo la proteina F, che ha potere fusogeno, e nel caso del virus del morbillo abbiamo anche la proteina H, che in questo schema si può vedere che è siglata come HN in quanto in alcuni paramixovirus, ma non nel virus del morbillo, questa proteina (H) ha associata anche l’attività di neuroamminidasi; nel virus del morbillo c’è invece solo una emagglutinina, glicoproteina che funge da anti-recettore, nel mediare l’incontro del virus con il recettore situato sulla cellula ospite.
Sempre a livello dell’envelope è presente una piccola proteina idrofobica (SH) della quale però non conosciamo ancora la funzione.
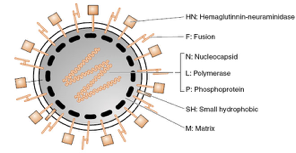
Al di sotto dell’envelope si trova la matrice (evidenziata con trattini neri nell’immagine) che contiene il genoma: come il virus dell’influenza, anche il virus del morbillo ha un genoma costituito da RNA a polarità negativa, ma a differenza del virus dell’influenza, è formato da un unico segmento di RNA lineare racchiuso nel capside, costituito da proteine capsidiche che creano una geometria elicoidale. Ovviamente, questo virus, possedendo un genoma a RNA a polarità negativa, deve contenere una RNA polimerasi RNA dipendente, che consenta a questo genoma, una volta arrivato nel citoplasma della cellula ospite, di essere espresso. Senza questa attività enzimatica, rappresentata nel caso del virus del morbillo dalle proteine L e P, il virus non può esprimersi, perché la cellula infetta non gli può mettere a disposizione una RNA polimerasi RNA dipendente. Al centro abbiamo il nucleocapside, costituito dalla proteina N, mentre le proteine L e P costituiscono insieme l’RNA polimerasi RNA dipendente.
Il ciclo replicativo del virus del morbillo
Nella diapositiva successiva sono mostrati in maggiore dettaglio le proteine localizzate nell’envelope del virus del morbillo, la glicoproteina F, dotata di potere fusogeno, e la glicoproteina H, che costituisce l’antirecettore.
Esaminando la glicoproteina F possiamo osservare una nuova modalità di ingresso dei virus nelle cellule; parlando di rinovirus e di influenza virus abbiamo visto che questi 2 entrano nella cellula attraverso un processo di endocitosi, il virus si ancora al recettore presente nella membrana citoplasmatica, una volta entrato, un pezzo di membrana si ripiega all’interno della cellula finché non si distacca nella vescicola endocitotica e successivamente, nel processo di uncoating, consentirà al genoma virale di essere liberato nel citoplasma. Il poliovirus invece, prende contatto col recettore, la proteina VP4, quella localizzata più esternamente nel capside del polio, salta, cambio conformazionale della proteina VP1, che si introflette, muovendo le sue estremità N-terminali, forma un poro e attraverso questo inietta il genoma del poliovirus.
Esaminando la glicoproteina F possiamo osservare una nuova modalità di ingresso dei virus nelle cellule; parlando di rinovirus e di influenza virus abbiamo visto che questi 2 entrano nella cellula attraverso un processo di endocitosi, il virus si ancora al recettore presente nella membrana citoplasmatica, una volta entrato, un pezzo di membrana si ripiega all’interno della cellula finché non si distacca nella vescicola endocitotica e successivamente, nel processo di uncoating, consentirà al genoma virale di essere liberato nel citoplasma. Il poliovirus invece, prende contatto col recettore, la proteina VP4, quella localizzata più esternamente nel capside del polio, salta, cambio conformazionale della proteina VP1, che si introflette, muovendo le sue estremità N-terminali, forma un poro e attraverso questo inietta il genoma del poliovirus.
Diversa modalità presenta invece il virus del morbillo, per la presenza di questo polipeptide endogeno presente nella superficie; la proteina F è divisa in 2 porzioni: la porzione maggiore e quella ancorata con l’estremità carbossi-terminale nella membrana cellulare e tramite un ponte disolfuro è legata ad una porzione più piccola che protrude verso l’esterno.
Cosa succede quando questa proteina F viene messa a contatto con la cellula ospite? Il contatto è stato realizzato dalla proteina H, che ha un cambio di conformazione, in seguito al quale la porzione fusogena viene esposta, e a pH neutro (quindi diversamente da quanto visto nelle proteine lisogene che lavoravano all’interno dei lisosomi) fa si che l’envelope virale possa fondersi direttamente a pH neutro, con la membrana citoplasmatica della cellula ospite, consentendo al nucleocapside di penetrare all’interno della cellula.
Ci sono virus che hanno genomi che vanno da 15 a 18 Kbp, ed una caratteristica dei genomi dei paramixovirus è di avere sempre una sequenza di geni estremamente ordinata e questo consente al virus di produrre maggiori quantità di proteine che più gli servono (ad es. proteine strutturali) e minori quantità di proteine meno necessarie (ad es. proteine enzimatiche, che possono essere usate più volte).
La glicoproteina presente nella superficie ha la possibilità di legarsi a 2 recettori: il primo recettore è rappresentato da molecole di CD46, molecola diffusa in moltissime cellule del nostro organismo, il che ci fa capire come, differentemente dal virus del raffreddore e dal virus dell’influenza, il virus del morbillo riesce a stabilire un’infezione sistemica, invece il virus del raffreddore, trasmesso per via aerea, si ferma nelle prime vie respiratorie (cavità nasali), dove trova una temperatura non molto elevata, di circa 34°C, ottima per la sua replicazione, pronto intervento delle difese di tipo innato, risoluzione dell’infezione e non si sposta da questa localizzazione anatomica; il virus dell’influenza invece si ferma nel tratto superiore delle vie respiratorie e rimane localizzato in questo sito.
Ci sono virus che hanno genomi che vanno da 15 a 18 Kbp, ed una caratteristica dei genomi dei paramixovirus è di avere sempre una sequenza di geni estremamente ordinata e questo consente al virus di produrre maggiori quantità di proteine che più gli servono (ad es. proteine strutturali) e minori quantità di proteine meno necessarie (ad es. proteine enzimatiche, che possono essere usate più volte).
La glicoproteina presente nella superficie ha la possibilità di legarsi a 2 recettori: il primo recettore è rappresentato da molecole di CD46, molecola diffusa in moltissime cellule del nostro organismo, il che ci fa capire come, differentemente dal virus del raffreddore e dal virus dell’influenza, il virus del morbillo riesce a stabilire un’infezione sistemica, invece il virus del raffreddore, trasmesso per via aerea, si ferma nelle prime vie respiratorie (cavità nasali), dove trova una temperatura non molto elevata, di circa 34°C, ottima per la sua replicazione, pronto intervento delle difese di tipo innato, risoluzione dell’infezione e non si sposta da questa localizzazione anatomica; il virus dell’influenza invece si ferma nel tratto superiore delle vie respiratorie e rimane localizzato in questo sito.
Diversa la situazione per il virus del morbillo, anch’esso trasmesso per via aerea, ma riesce a spostarsi, perché all’inizio si localizza nel tratto superiore delle vie respiratorie e poi trova il modo per realizzare un’infezione sistemica. Di fatto il recettore CD46 è presente in molti tipi di cellule del nostro organismo, la cui funzione fisiologica è proteggere le nostre cellule dall’attacco di componenti del complemento, quindi ha un’attività difensiva nei riguardi del complemento. Ma il virus del morbillo non si limita a riconoscere quest’unico recettore, ma anche un secondo, rappresentato da CD150/SLAM , una molecola importante nell’attivazione del linfociti T nel corso della risposta immunitaria, e questo CD150 si trova espresso sui linfociti B, linfociti T e sulle cellule dendritiche, attaccando e replicando in queste cellule, che sono elementi cardine nello svolgimento della risposta immunitaria, perciò questo virus è in grado di indurre immunosoppressione.
Nei Paesi sviluppati, grazie ad un vaccino, il morbillo è oggi un’infezione controllata, mentre nei Paesi sottosviluppati ci sono ogni anno milioni di casi di infezione e un milione di morti, soprattutto bambini, a causa del morbillo, soprattutto perché non è possibile praticare la vaccinazione e rimane un grosso problema a livello sanitario.
Riprendendo il ciclo replicativo, la proteina F che forma l’envelope lisa la membrana citoplasmatica, proteina H che riconosce il recettore CD46 o il recettore CD150, quindi ingresso diretto del nucleocapside nel citoplasma. Il genoma, a polarità negativa, a questo punto deve essere trascritto dall’RNA polimerasi RNA dipendente perché il virus possa esprimersi.
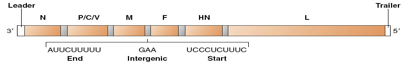
Soffermiamoci un attimo ad analizzare questo genoma: all'estremità 3' è presente una sequenza leader, importante perché presenta un sito d'attacco per l'RNA polimerasi RNA dipendente, come pure è importante per dare inizio al processo di incapsidamento, cioè quando il genoma viene rivestito dalle proteine strutturali che assumono poi simmetria elicoidale. Seguono poi, con un ordine conservato, la proteina N (proteina strutturale), la proteina P (cofattore dell'RNA pol RNA dip) (nell'immagine c'è “P/C/V” perché dallo stesso segmento genico, oltre che la proteina P, vengono prodotte anche la proteina C e la proteina V), segue poi il gene che porta alla sintesi della proteina di matrice, il gene della proteina fusogena localizzata nell'envelope, il gene che sintetizza l'emoagglutinina, perché non c'è una neuroaminidasi nel virus del morbillo, e ultima, la proteina L, che è l'RNA polimerasi RNA dipendente.
La successione di questi geni viene conservata nei paramixovirus.
Tra un gene e l'altro sono presenti delle sequenze intergeniche che sono molto importanti per capire come questo genoma a polarità negativa viene trascritto per dare origine agli mRNA. Ci sono 2 modelli che descrivono l'azione dell'RNA pol RNA dip: il primo dice che l'enzima riconosce i promotori presenti a monte dei rispettivi geni, il secondo, che è poi quello confermato, afferma che l'RNA pol non riconosce promotori a monte di ciascun gene, ma si lega solo alla sequenza leader in posizione 3' e procede trascrivendo il genoma fino all'estremità 5'. Si è arrivati a stabilire questo con un semplice esperimento: irradiando il genoma con UV si formano dei dimeri di uridina, quando la polimerasi scorre e incontra questi dimeri smette di trascrivere e si stacca dallo stampo; se esistessero molti promotori, la mutazione avrebbe effetto solo sulla sintesi di un gene, mentre gli altri mRNA verrebbero trascritti in modo indipendente; invece, se il promotore è presente solo nella sequenza leader, in presenza di una mutazione, si avrebbe la trascrizione di tutti i geni presenti a monte del dimero di uridina, mentre i geni a valle non potrebbero essere trascritti (che è proprio quello che succede, dimostrando che esiste un unico punto di legame della polimerasi al genoma).
Fino ad ora abbiamo analizzato come avviene la sintesi dell’mRNA, il problema è che questo RNA, che presenta spazi intergenici, deve anche essere copiato in un anti-genoma, il genoma negativo, cioè a polarità positiva, che a sua volta deve fungere da stampo per nuovi genomi del virus del morbillo; per questo è necessario qualche cambiamento. Il meccanismo che determina il cambiamento dell’RNA pol, affinché smetta di trascrivere messaggeri e cominci a copiare l’intero genoma, per avere poi stampi indispensabili per la formazione di nuovi genomi pronti a partecipare al processo di morfogenesi del virus del morbillo all’interno della cellula; le cose vanno in questa maniera: il genoma a polarità negativa, con i suoi spazi intergenici importanti nel regolare la trascrizione, funge da stampo per la sintesi di un genoma a polarità positiva, questo è possibile quando c’è molta proteina capsidica che “tappa”, nasconde quegli spazi intergenici indispensabili per la trascrizione; per fungere da stampo per la formazione di anti-genomi positivi è necessario che sia rivestito di proteina capsidica, devono avvenire 2 eventi in maniera sequenziale: dal genoma a polarità negativa si deve avere una produzione di una progenie della stessa lunghezza del genoma, però a polarità positiva, che serve da stampo per la formazione di nuovi genomi. Ma qual è il meccanismo di controllo che dice “stop alla sintesi dei mRNA” perché ci sono abbastanza proteine per la formazione di matrice, capsidi, ecc. a questo punto il virus si deve preoccupare di inserire nei capsidi dei nuovi genomi e avviene in questo modo (vedi slide):
Altra strategia che il virus impiega è determinata dal fatto che, come abbiamo visto precedentemente, quando sta preparandosi per il processo di gemmazione espone la proteina H e la proteina F sulla propria superficie: questa è una cellula infetta e la proteina F così esposta si lega ad una cellula sana vicina e il virus si trasmette da una cellula all'altra senza passare in ambiente extracellulare, si formano delle “cellule giganti”, si apre la possibilità per i nucleocapsidi di passare da una cellula all'altra e in questo modo il virus non si espone agli anticorpi circolanti, riuscendo a fuggire dalla risposta umorale. Quindi altra strategia grazie alla presenza di proteine esogene il virus forma queste “cellule giganti” e multinucleate che lo proteggono dall'incontro con gli anticorpi.
Abbiamo a disposizione un vaccino, si tratta di un vaccino attenuato, perché sottoposto a modifiche tali da rendere incapaci i virus di indurre la malattia: in altre parole, la virulenza è attenuata e il virus non è in grado di stimolare la produzione di anticorpi; viene somministrato come vaccino trivalente (morbillo-rosolia-parotite) e viene somministrato per via sottocutanea nella parte esterna del braccio.
Genoma codifica per 6 geni, in ordine 3´N-P/C/V-M-F-H-L5´.
Ciascuna giunzione fra i geni contiene una sequenza consenso che include
> il sito di termine della trascrizione di un gene
> Il sito di poliadenilazione
> due-tre nucleotidi intergenici
> il sito di inizio della trascrizione del gene sucecssivo
Ciascuna giunzione fra i geni contiene una sequenza consenso che include
> il sito di termine della trascrizione di un gene
> Il sito di poliadenilazione
> due-tre nucleotidi intergenici
> il sito di inizio della trascrizione del gene sucecssivo
I geni che vengono codificati sono 6: N, P, M, F, H e L; l’RNA polimerasi trascrive gli mRNA, dotandoli di una coda di poliA all’estremità 3’, questo è possibile grazie alla sequenza intergenica presente tra un gene e l’altro, che contiene una sequenza consenso per il sito di termine per la trascrizione di un gene, il sito di poliadenilazione, 2-3 nucleotidi che dividono la sintesi di un gene da quello successivo e infine il sito di inizio della trascrizione del gene successivo.
Questi mRNA però non sono sintetizzati tutti nella stessa quantità (più N ed M, ma meno L, ad esempio) e la realizzazione di questo dipende dal fatto che quando arriva una porzione intergenica, la possibilità che vada avanti l’RNA pol a trascrivere il gene successivo diminuisce dal 60 all’80%, quindi c’è un 20% di probabilità che vada avanti; questo comporta una produzione di grandi quantità di N, poi una minor quantità di P, di M e così via, fino ad arrivare a L che ha attività enzimatica e quindi ne servono meno copie; in questo modo il virus effettua un controllo quantitativo delle proteine che gli servono; questo fenomeno è noto come ATTENUAZIONE DELLA TRASCRIZIONE ed ha ovviamente come conseguenza la sintesi di messaggeri, alcuni dei quali prodotti in abbondanza, altri in minor quantità. La quantità di prodotto genico virale è inversamente proporzionale alla distanza del gene dal 3’ dello stampo, quanto è più vicino al 3’ (come N) tanto più quella proteina sarà prodotta, tanto più sarà distante (come L) tanto minore sarà la quantità di quella proteina prodotta.
In più, non c’è un esatta corrispondenza mRNA-proteina, ad esempio, il gene P da origine non solo alla proteina P, cofattore dell’RNA pol, ma anche ad altre 2 proteine: C e V. Questo è possibile grazie ad un processo definito “P-RNA editing”; in questo gene abbiamo una sequenza di poliU, quando l’RNA pol copia questa sequenza, per complementarietà si avrà una sequenza di poliA, il che comporta un rallentamento e poi blocco dell’RNA pol, che dopo aver fatto una pausa torna indietro, si ha una sorta di lettura falsata (nell’esempio una G viene letta 2 volte, quindi se ne avranno 4 invece di 3) e questi nuovi moduli che si sono formati, incontrando precocemente rispetto al gene P un segnale di stop, portano alla sintesi di altre 2 proteine.
In più, non c’è un esatta corrispondenza mRNA-proteina, ad esempio, il gene P da origine non solo alla proteina P, cofattore dell’RNA pol, ma anche ad altre 2 proteine: C e V. Questo è possibile grazie ad un processo definito “P-RNA editing”; in questo gene abbiamo una sequenza di poliU, quando l’RNA pol copia questa sequenza, per complementarietà si avrà una sequenza di poliA, il che comporta un rallentamento e poi blocco dell’RNA pol, che dopo aver fatto una pausa torna indietro, si ha una sorta di lettura falsata (nell’esempio una G viene letta 2 volte, quindi se ne avranno 4 invece di 3) e questi nuovi moduli che si sono formati, incontrando precocemente rispetto al gene P un segnale di stop, portano alla sintesi di altre 2 proteine.
Questo è un esempio di come i virus, non avendo una grande quantità di informazioni, hanno evoluto dei meccanismi che gli consentono di sfruttare al massimo il proprio genoma. Le proteine V e C sono importanti nel difendere il virus dagli attacchi delle difese immunitarie dell’ospite.
Fino ad ora abbiamo analizzato come avviene la sintesi dell’mRNA, il problema è che questo RNA, che presenta spazi intergenici, deve anche essere copiato in un anti-genoma, il genoma negativo, cioè a polarità positiva, che a sua volta deve fungere da stampo per nuovi genomi del virus del morbillo; per questo è necessario qualche cambiamento. Il meccanismo che determina il cambiamento dell’RNA pol, affinché smetta di trascrivere messaggeri e cominci a copiare l’intero genoma, per avere poi stampi indispensabili per la formazione di nuovi genomi pronti a partecipare al processo di morfogenesi del virus del morbillo all’interno della cellula; le cose vanno in questa maniera: il genoma a polarità negativa, con i suoi spazi intergenici importanti nel regolare la trascrizione, funge da stampo per la sintesi di un genoma a polarità positiva, questo è possibile quando c’è molta proteina capsidica che “tappa”, nasconde quegli spazi intergenici indispensabili per la trascrizione; per fungere da stampo per la formazione di anti-genomi positivi è necessario che sia rivestito di proteina capsidica, devono avvenire 2 eventi in maniera sequenziale: dal genoma a polarità negativa si deve avere una produzione di una progenie della stessa lunghezza del genoma, però a polarità positiva, che serve da stampo per la formazione di nuovi genomi. Ma qual è il meccanismo di controllo che dice “stop alla sintesi dei mRNA” perché ci sono abbastanza proteine per la formazione di matrice, capsidi, ecc. a questo punto il virus si deve preoccupare di inserire nei capsidi dei nuovi genomi e avviene in questo modo (vedi slide):
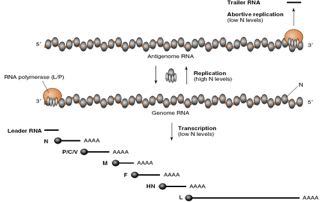
in posizione 3' dell'RNA genomico a polarità negativa è presente un'importante sequenza LEADER, la quale viene riconosciuta dal promotore dell'RNA polimerasi e dà l'avvio al processo di trascrizione, ma è molto importante anche per stabilire il momento in cui il virus deve iniziare a sintetizzare nuovi genomi. Molto importante per questa funzione è anche la sequenza TRAILER che troviamo in posizione 3' dell'antigenoma a polarità positiva, copiato dal genoma. Queste sequenze sono importanti perché il virus sarà pronto a fabbricare nuovi genomi solo quando avrà a disposizione molte proteine N (nucleocapsidiche), le quali si legano alla sequenza LEADER del genoma e lo trascrivono completamente, senza nessun salto tra un gene e l'altro, portando alla formazione dell'antigenoma; quest'ultimo, sempre in presenza di grandi quantità di N, funge così da stampo per la sintesi di nuovi genomi.
Quando le proteine N sono poche, invece, non è necessaria la sintesi di nuovi genomi e queste interagiscono sia con la sequenza LEADER che con la sequenza TRAILER: la polimerasi si lega, può trascrivere la regione LEADER dell'RNA, un grosso frammento viene rilasciato e la polimerasi, secondo lo schema analizzato prima, trascrive tutti i messaggeri che servono per la produzione delle proteine, indispensabili al virus per la replicazione.
Quindi, solo quando la concentrazione di N aumenta, la sequenza LEADER viene rivestita dalle proteine capsidiche e la molecola genomica verrà copiata in un antigenoma. Anche nell'antigenoma però deve esserci un segnale che dice “di antigenomi non ne servono molti perché ci sono poche proteine strutturali”, perciò la sequenza TRAILER in posizione 3', quando ci sono poche proteine N, trascrive un corto frammento e dopo di che la polimerasi, non essendoci altri siti di inizio, si distacca e si realizza quello che viene chiamato un processo replicativo abortivo.
Riassunto: quando il virus è pronto ad impacchettare nuovi genomi deve essere prodotta una grande quantità di proteine, alcune delle quali si legano alla molecola di RNA a polarità negativa, quindi il genoma, e mascherano i segni intergenici, che hanno il compito di guidare la polimerasi mentre sta trascrivendo per rispettare determinati segnali di stop; come risultato di questo mascheramento, la polimerasi è capace di produrre una catena di RNA positiva completa, che a sua volta può essere copiata e dar luogo alla formazione di genomi a polarità negativa.
Abbiamo suggerito che una sequenza leader è presente in posizione 3' del genoma e una sequenza trailer è presente in posizione 3' dell'antigenoma: se ci sono poche proteine N, per dare inizio al processo di morfogenesi, la polimerasi trascrive la sequenza leader e riconosce i segnali di stop nelle sequenze intergeniche, portando così alla trascrizione di messaggeri.
Il controllo a livello dell'antigenoma: quando le proteine strutturali sono tante la polimerasi trascrive per formare nuovi genomi, quando le proteine N sono poche la polimerasi trascrive la sequenza trailer, si ferma, la replicazione è abortiva e non vengono, di conseguenza, formati nuovi antigenomi.
Questa figura, in modo molto schematico, riassume il ciclo di replicazione dei paramixovirus:
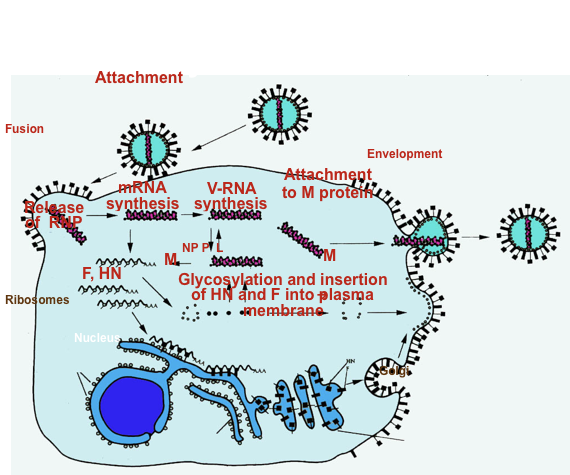
l'attacco, mediato da 2 diversi recettori, uno dei quali presente su linfociti T, B e cellule dendritiche, l'altro (CD46) diffuso in molte cellule del nostro organismo, grazie alla presenza di una proteina fusogena che può funzionare a pH neutro, l'envelope del virus si fonde con la membrana citoplasmatica della cellula e vengono rilasciate le ribonucleoproteine. La prima sintesi macromolecolare virus-specifica è un processo di trascrizione realizzato dall'RNA polimerasi RNA dipendente che il virus si porta dietro; si formano gli mRNA, che daranno origine alle proteine di matrice, alle proteine nucleocapsidiche, alle proteine P ed L che insieme formano la polimerasi, le proteine di superficie H ed F, che grazie al passaggio nel RE e nel Golgi verranno inserite nella membrana citoplasmatica della cellula all'interno della quale il virus del morbillo sta replicando.
Queste zone dove le glicoproteine sono state inserite vengono rivestite dalla proteina N, nucleocapsidi in neoformazione, probabilmente veicolati dalla proteina M, raggiungono dei punti nella membrana citoplasmatica della cellula e il virus fuoriesce per un processo di gemmazione.
La risposta immunitaria al virus del morbillo
Analogamente a raffreddore ed influenza, anche il morbillo è trasmesso con starnuti, colpi di tosse e ambienti poco areati sono ideali per questo tipo di trasmissione. Però c'è una grossa differenza di comportamento tra il virus dell'influenza e il virus del morbillo: mentre il primo entra nelle vie respiratorie e rimane localizzato lì, il secondo ha un comportamento molto differente.
Entra nell'organismo attraverso le prime vie respiratorie, trova nelle cellule il recettore CD46 e riesce ad attaccarsi e a replicare; non si ha una grossa produzione di virus perché c'è contemporaneamente una difesa immunitaria innata abbastanza sostenuta che controlla questa infezione; però a questo punto sono presenti nel sito di infezione delle cellule dendritiche: qual è il loro compito? Spostarsi dal sito dell'infezione al linfonodo, perché possa essere attivata l'immunità di tipo adattativo.
Quando nelle vie respiratorie c'è un virus influenzale, le cellule dendritiche catturano il virus e presentano pezzi di antigene nel linfonodo per allertare i linfociti T; ma la differenza fondamentale tra questi 2 tipi di infezione, che apre la porta al morbillo all'infezione sistemica, è che grazie alle cellule dendritiche il virus dell'influenza viene semplicemente trasportato all'interno del linfonodo, mentre il virus del morbillo si replica al loro interno, spostando perciò l'infezione dall'apparato respiratorio al linfonodo.
Entra nell'organismo attraverso le prime vie respiratorie, trova nelle cellule il recettore CD46 e riesce ad attaccarsi e a replicare; non si ha una grossa produzione di virus perché c'è contemporaneamente una difesa immunitaria innata abbastanza sostenuta che controlla questa infezione; però a questo punto sono presenti nel sito di infezione delle cellule dendritiche: qual è il loro compito? Spostarsi dal sito dell'infezione al linfonodo, perché possa essere attivata l'immunità di tipo adattativo.
Quando nelle vie respiratorie c'è un virus influenzale, le cellule dendritiche catturano il virus e presentano pezzi di antigene nel linfonodo per allertare i linfociti T; ma la differenza fondamentale tra questi 2 tipi di infezione, che apre la porta al morbillo all'infezione sistemica, è che grazie alle cellule dendritiche il virus dell'influenza viene semplicemente trasportato all'interno del linfonodo, mentre il virus del morbillo si replica al loro interno, spostando perciò l'infezione dall'apparato respiratorio al linfonodo.
Abbiamo visto che ci sono cellule come macrofagi, linfociti T e B che presentano il secondo recettore che il virus può utilizzare, ed ecco che il linfonodo si trasforma in un centro di infezione, dal linfonodo raggiunge il sistema ematico e si ha di conseguenza un'infezione sistemica. Cellule CD46 sono diffuse in tutto il nostro organismo ed ecco che, proprio per questa disseminazione per via ematica, il virus del morbillo torna, ma in numero di molto superiore rispetto all'iniziale infezione, a riprodursi a livello dell'apparato respiratorio; ci deve essere una forte risposta immunitaria adattativa a questo punto per risolvere l'infezione, ed è questo il momento in cui si può facilmente trasmettere l'infezione ad altri soggetti. La chiave di tutto è la capacità di questo virus di aprirsi le porte ad un'infezione sistemica riuscendo a replicare a livello delle cellule dendritiche.
Il virus del morbillo “fugge” dai confini del tratto respiratorio, entra nel sistema linfatico e stabilisce un'infezione sistemica; dopo di che, attacca per una seconda volta le vie respiratorie, stavolta attaccando e uccidendo un numero elevato di cellule e può infine essere trasmesso ad altri individui.
Stiamo parlando di un virus ad RNA ed una caratteristica che accomuna tutti i virus che hanno questo tipo di genoma e che riguarda l'enzima che ne guida la trascrizione è il fatto di possedere una RNA polimerasi RNA dipendente, che però non possiede un'elevata fedeltà, di conseguenza tutti gli RNA virus vanno incontro a quella che viene chiamata DERIVA ANTIGENICA. La polimerasi funziona male e crea delle mutazioni, le quali si possono ripercuotere sulla struttura superficiale del virus, alterando per es. il suo recettore, come nel caso del virus dell'influenza, dove in seguito a queste mutazioni l'emoagglutinina cambia e non è più neutralizzata da anticorpi precedentemente formati; va incontro a deriva antigenica anche il virus del raffreddore, e tutti i virus che possiedono questo tipo di polimerasi, la quale va incontro ad errore a volte anche solo ogni 1000 bp. La deriva antigenica porta alla circolazione di un centinaio di sierotipi differenti di rinovirus.
Il virus del morbillo “fugge” dai confini del tratto respiratorio, entra nel sistema linfatico e stabilisce un'infezione sistemica; dopo di che, attacca per una seconda volta le vie respiratorie, stavolta attaccando e uccidendo un numero elevato di cellule e può infine essere trasmesso ad altri individui.
Stiamo parlando di un virus ad RNA ed una caratteristica che accomuna tutti i virus che hanno questo tipo di genoma e che riguarda l'enzima che ne guida la trascrizione è il fatto di possedere una RNA polimerasi RNA dipendente, che però non possiede un'elevata fedeltà, di conseguenza tutti gli RNA virus vanno incontro a quella che viene chiamata DERIVA ANTIGENICA. La polimerasi funziona male e crea delle mutazioni, le quali si possono ripercuotere sulla struttura superficiale del virus, alterando per es. il suo recettore, come nel caso del virus dell'influenza, dove in seguito a queste mutazioni l'emoagglutinina cambia e non è più neutralizzata da anticorpi precedentemente formati; va incontro a deriva antigenica anche il virus del raffreddore, e tutti i virus che possiedono questo tipo di polimerasi, la quale va incontro ad errore a volte anche solo ogni 1000 bp. La deriva antigenica porta alla circolazione di un centinaio di sierotipi differenti di rinovirus.
Il CAMBIAMENTO ANTIGENICO invece, è una diversa modalità che porta alla formazione di nuove particelle virali e trova la base nel rimescolamento di virus di specie diverse, che hanno un genoma segmentato, e quindi si possono scambiare tra loro dei pezzi, portando alla formazione di virus, ad es. dell'influenza, che in superficie hanno non un'emoagglutinina leggermente modificata, come nel caso della deriva antigenica, ma una completamente nuova.
Salta fuori un virus umano che presenta sulla sua superficie un'emoagglutinina che ha “fregato” ad un virus aviario e ovviamente nessuno in tutto il globo possiede anticorpi per quella proteina e si può scatenare di conseguenza una pandemia.
Quindi si può dire che, possedendo il virus del morbillo una RNA polimerasi RNA dipendente, sicuramente va incontro ad una deriva antigenica, ma non può andare incontro ad un cambiamento antigenico perché il suo genoma non è segmentato; infatti, noi ci ammaliamo una volta sola e acquistiamo un'immunità che dura per tutta la vita. Questo perché nonostante la polimerasi faccia numerosi errori durante la replicazione non ha nessuna ripercussione sugli effetti patogeni del virus; questo dipende dal fatto che il recettore responsabile del legame del virus alla cellula non ammette mutazioni, se insorge una mutazione che abbia ripercussioni su questo recettore, il recettore perde la capacità di legarsi al recettore cellulare e non può avere inizio il ciclo di replicazione virale.
Perciò abbiamo un unico sierotipo, anche se si formeranno diversi virus con una superficie alterata, ma questa superficie alterata non può svolgere la propria funzione, e fanno parte di virus difettivi che andranno ben presto ad essere eliminati e questo, infine, spiega perché esiste un unico sierotipo, l'unico ospite naturale è l'uomo e le mutazioni fanno sì che solo un sierotipo sia in circolazione. Questa può essere considerata una prima difesa, in quanto lo aiuta a non farsi selezionare dalla produzione di tanti tipi diversi di anticorpi e crea anche un problema, che ha consentito di capire che è un virus (in termini virologici 6000 anni fa non sono molti) che si ritiene essere nato abbastanza recentemente; questo dipende dal fatto che è un virus estremamente infettante, infatti quando in una popolazione circola il morbillo, tutti gli individui suscettibili, che non hanno contratto il virus precedentemente si ammalano. Non potendo andare incontro a deriva antigenica, deve crearsi la possibilità di instaurare una catena di trasmissione ininterrotta: lui non può mutare, infettandolo tutti quanti (?) metterebbe a rischio la propria sopravvivenza; ecco che il virus si è potuto diffondere per la prima volta sulla faccia della Terra quando gli individui hanno iniziato a vivere insieme.
Perciò abbiamo un unico sierotipo, anche se si formeranno diversi virus con una superficie alterata, ma questa superficie alterata non può svolgere la propria funzione, e fanno parte di virus difettivi che andranno ben presto ad essere eliminati e questo, infine, spiega perché esiste un unico sierotipo, l'unico ospite naturale è l'uomo e le mutazioni fanno sì che solo un sierotipo sia in circolazione. Questa può essere considerata una prima difesa, in quanto lo aiuta a non farsi selezionare dalla produzione di tanti tipi diversi di anticorpi e crea anche un problema, che ha consentito di capire che è un virus (in termini virologici 6000 anni fa non sono molti) che si ritiene essere nato abbastanza recentemente; questo dipende dal fatto che è un virus estremamente infettante, infatti quando in una popolazione circola il morbillo, tutti gli individui suscettibili, che non hanno contratto il virus precedentemente si ammalano. Non potendo andare incontro a deriva antigenica, deve crearsi la possibilità di instaurare una catena di trasmissione ininterrotta: lui non può mutare, infettandolo tutti quanti (?) metterebbe a rischio la propria sopravvivenza; ecco che il virus si è potuto diffondere per la prima volta sulla faccia della Terra quando gli individui hanno iniziato a vivere insieme.
Altra strategia che il virus impiega è determinata dal fatto che, come abbiamo visto precedentemente, quando sta preparandosi per il processo di gemmazione espone la proteina H e la proteina F sulla propria superficie: questa è una cellula infetta e la proteina F così esposta si lega ad una cellula sana vicina e il virus si trasmette da una cellula all'altra senza passare in ambiente extracellulare, si formano delle “cellule giganti”, si apre la possibilità per i nucleocapsidi di passare da una cellula all'altra e in questo modo il virus non si espone agli anticorpi circolanti, riuscendo a fuggire dalla risposta umorale. Quindi altra strategia grazie alla presenza di proteine esogene il virus forma queste “cellule giganti” e multinucleate che lo proteggono dall'incontro con gli anticorpi.
Altra differenza è che questo virus induce immunosoppressione, per il fatto che varie cellule del sistema immunitario (linfociti T e B, macrofagi, ecc) possiedono un recettore riconosciuto dal virus e vengono perciò infettate e uccise; nei Paesi sottosviluppati è ancora altissimo il numero di morti annuali in seguito all'infezione del morbillo e molto spesso la morte di questi individui è dovuta ad una polmonite, perché l'epitelio respiratorio è già danneggiato dall'azione dei virus, i batteri perciò si impiantano facilmente, si ha un'immunodepressione, perciò l'individuo non risponde efficacemente all'infezione batterica e si ha sviluppo di polmonite, che è una delle principali cause di morte in questi casi.
Caratteristico di questa malattia è il rash cutaneo provocato da questa malattia ed insorge perché, dopo essersi diffuso per via ematica, il virus attacca le cellule endoteliali e poi passa nell’epitelio, inducendo così la produzione di citochine da parte di cellule che cercano di contrastare quest’infezione.
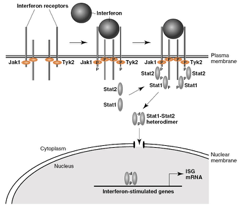
Un’altra arma di difesa da parte dell’organismo è la produzione di interferone, anche se l’induzione da parte del virus del morbillo non è così elevata come per il virus dell’influenza; si ritiene che le proteine C e V (generate dal gene P) costituiscano un fattore di virulenza, in quanto modulano e interferiscono con il potere antivirale dell’interferone e queste 2 proteine in particolare portano alla degradazione delle proteine stat1 e stat2: l’interferone viene prodotto all’interno della cellula infettata, il punto di partenza è solitamente una molecola di RNA bicatenario, induzione di fattori di trascrizione che si complessano nel sito dell’enhancer dell’interferone, produzione dell’interferone, che viene portato in ambiente extracellulare e va ad allertare le cellule vicine. L’interferone comunica tra una cellula e l’altra attraverso dei recettori (jak1 e tyk2), che lo legano e in seguito vengono fosforilati e si avvicinano, essendo delle kinasi, e a questo punto l’informazione deve raggiungere il nucleo perché vengano prodotte proteine in grado di contrastare l’interferone. Le proteine C e V bloccano le proteine stat1 e stat2 che, dopo esser state fosforilate, hanno il compito determinante di portare il segnale nel nucleo, in maniera tale che i geni stimolati dall’interferone possano essere prima trascritti e poi tradotti.
Abbiamo a disposizione un vaccino, si tratta di un vaccino attenuato, perché sottoposto a modifiche tali da rendere incapaci i virus di indurre la malattia: in altre parole, la virulenza è attenuata e il virus non è in grado di stimolare la produzione di anticorpi; viene somministrato come vaccino trivalente (morbillo-rosolia-parotite) e viene somministrato per via sottocutanea nella parte esterna del braccio.
Continua a leggere:
- Successivo: I virus a trasmissione orale: l'epatite A
- Precedente: Il Rinovirus
Dettagli appunto:
-
Autore:
Simone Pisu
[Visita la sua tesi: "Caratterizzazione molecolare ed eterogenicità delle varianti emoglobiniche in Sardegna"]
- Università: Università degli Studi di Cagliari
- Facoltà: Biologia
- Corso: Biologia Cellulare e Molecolare
- Esame: Virologia molecolare
- Docente: Prof. Marongiu
Altri appunti correlati:
- Microbiologia clinica
- Biologia applicata
- Microbiologia
- Microbiologia Medica
- Filologia della letteratura italiana
Per approfondire questo argomento, consulta le Tesi:
- Piante medicinali come ausilio per contrastare le patologie virali: tre casi-studio
- Contributo allo studio ed alla terapia dell'epatite C nel paziente HIV-positivo
- Memetica e nuovi media
- Peptidi overlapping derivanti da sequenze gH-omologhe di HSV-1 come inibitori dell'attività virale.
- Selezione di Mutanti HIV-1 Resistenti e Basi Molecolari della Resistenza di Nuovi Inibitori Non-Nucleosidici della Trascrittasi Inversa
Puoi scaricare gratuitamente questo appunto in versione integrale.
 Ricevi informazioni sui nostri servizi, sulle offerte e non perdere news e consigli su università e lavoro.
Ricevi informazioni sui nostri servizi, sulle offerte e non perdere news e consigli su università e lavoro.